Этиология.
Возбудители ИКБ относятся к порядку Spirochaetales, семейству Spirochaetaceae, роду Borrelia. Свое родовое название они получили в честь французского микробиолога A. Borrel, изучавшего в начале ХХ века эту разновидность спирохет.
В настоящее время по отличиям в нуклеотидных последовательностях ДНК различают более 18 геновидов боррелий, относящихся к комплексу B. burgdorferi sensu lato, представители которого неравномерно распределены в пределах мирового нозоареала этой инфекции. Не все боррелии данного комплекса являются патогенными для человека. Доказана патогенность 3 геновидов: B. burgdorferi sensu stricto, B. garinii, B. afzelii. Все эти геновиды имеют распространение на территории России (преимущественно B. garinii и B. afzelii). Еще 6 геновидов были выделены у человека: B. spielmanii, B. bavariensis, B. valaisiana, B. lusitaniae, B. bissetii, но их значение в развитии клинических синдромом пока окончательно не определено.
Это грамотрицательные спирохеты, относительно легко окрашиваются анилиновыми красителями, что отличает их от других родов спирохет. Другой отличительной особенностью боррелий является отсутствие у них митохондрий и ундулирующей мембраны. Длина микробной клетки - от 10 до 30 мкм, поперечный размер составляет 0,20-0,25 мкм. По форме представляют собой извитую, лево- или правовращающуюся спираль.
Боррелии имеют одну линейную хромосому и до 20 кольцевых и линейных плазмид, известных еще под названием - микрохромосомы. Белковый (антигенный) спектр боррелий весьма вариабелен и различен не только у боррелий разных геновидов, но и среди изолятов одного геновида.
Боррелии имеют группы антигенов: поверхностные (OspA, OspB, OspD, OspE и OspF), жгутиковый и цитоплазматический. Жгутиковый белок (флагеллин) массой 41 Кда имеет перекрестные детерминанты с основным белком миелина, белками кардиомиоцитов и синовиальной оболочки, а также с антигенами некоторых бактерий, что позволяет ему быть причиной как аутоимунных, так и ложноположительных реакций. Клеточная стенка представлена поверхностными липопротеинами (outer – surface protein): OspA, OspB, OspD, OspE и OspF, антигены которых очень изменчивы. Антигенный спектр боррелий вариабелен у различных штаммов одного геновида, а общность антигенной структуры боррелий между при их генотипической вариабельности составляет 48-70%. Поверхностные липопротеины определяют видовую принадлежность и являются основными иммуногенами. Наибольшая их гетерогенность обнаруживается у B. garinii и B. afzelii. Штаммы B.burgdorferi s.s. отличаются относительным постоянством спектра поверхностных белков. Антигенная схожесть боррелий с тканями макроорганизма (мимикрия) определяет развитие аутоиммунных процессов, имеющих наибольшую выраженность при хроническом течении. Многие антигенные детерминанты внешней оболочки сходны с таковыми у боррелий других видов и даже некоторых бактерий. Этим объясняется возможность возникновения перекрестных иммунологических реакций.
Молекулярно-генетические особенности возбудителя определяют полиморфизм клинических проявлений боррелиозной инфекции. Доказана относительная связь между геновидом боррелий и преимущественными органными поражениями. Поражение опорно-двигательного аппарата наиболее часто обусловлено геновидом B. burgdorferi sensu strico, неврологические поражения (нейроборрелиоз) ассоциирован с геновидом B. garinii, кожные поражения, в частности, ХААД наиболее часто регистрируются при инфицировании геновидом B. afzelii. При этом В. afzеlii и B. garinii обнаруживались в биоптатах из КМЭ [1].
Боррелии in vitro культивируются только в среде, обогащенной аминокислотами, витаминами, альбумином бычьей и кроличьей плазмы и другими веществами (модифицированная среда BSK-II). Температурный оптимум культивирования составляет 30-34оС. Боррелии отличаются относительно длительным и непостоянным периодом роста на питательных средах в анаэробных условиях, который обычно составляет 7-20 часов. При достижении числа микробных клеток 106 – 108 в мл возможно осуществление дальнейших лабораторных исследований. Боррелии, выращенные в среде культивирования, хорошо сохраняются при низких температурах (-70°- 90°С) до нескольких лет, не теряя своих биологических свойств. Формалин, фенол, этиловый спирт и другие дезинфицирующие вещества, а также ультрафиолетовое излучение обладают инактивирующим действием на боррелии.
Отсутствие у боррелий генов кодирования синтеза амино- и жирных кислот, ко-факторов и нуклеотидов объясняет столь высокие требования к условиям их культивирования с использованием сывороточных добавок. Кроме того, поскольку боррелии хемоорганотрофные анаэробы, то они должны получать энергию для синтеза АТФ от субстрата фосфорилирования, который имеется только в цитоплазме других клеток, т.к. у боррелий нет генов, кодирующих дыхательную цепь фосфорилирования. Боррелии также испытывают недостаток других ферментов, необходимых для цикла трикарбоновых кислот и фосфорилирования. Таким образом, способность их собственных компонентов метаболизма настолько ограничена, что для своего существования они должны иметь организм хозяина (причем с преимущественным внутриклеточным персистированием), который гарантирует доступность питательных готовых веществ. Кроме того, боррелии могут образовывать атипичные неподвижные L-формы (или псевдоцисты), для которых характерны пониженные обменные процессы, обеспечивающие возбудителю длительное выживание. При формировании псевдоцисты боррелии приобретают скрученную форму, при этом изменяются поверхностные белки, что позволяет возбудителю избегать иммунных реакций макроорганизма. Триггером образования псевдоцист является высокая температура, приемантибактериальных средств системного действия (в частности, бета-лактамных антибактериальных препаратов: пенициллинов). В последующем при благоприятных условиях среды псевдоцисты могут снова трансформироваться в подвижные спиралевидные формы. Способность образовывать L-формы является важным механизмом в развитии хронической инфекции [1, 3, 4, 5].
Патогенез.
Возбудители патогенного комплекса Borrelia burgdorferi sensu lato способны к диссеминации и длительной персистенции в организме человека, вследствие чего могут формироваться хронические формы заболевания, а также возможно цикличное и вариабельное течение инфекционного процесса в виде системного поражения органов-мишеней – кожи, сердца, нервной системы, опорно-двигательного аппарата [2, 3, 4, 5].
Боррелии обитают в эпителии кишечника голодного клеща. При кровососании боррелии размножаются, выходят в гемолимфу, в слюнные железы, и со слюной клеща попадают в кожу нового хозяина. Инфицирующие спирохеты экспрессируют декоринсвязывающий белок, посредством которого прикрепляются к протеогликанам. Не имеющие собственных протеаз, спирохеты утилизируют плазминоген-активаторную систему макроорганизма для деградации богатого фибриногеном экстрацеллюлярного матрикса. Это дает возможность микроорганизму «расплавлять» окружающие ткани, распространяться в коже и проникать в сосуды. Связывая неиммуногенные фибринолитические протеазы, боррелии реализуют новый механизм микробного патологического действия, который объясняет их способность к быстрой диссеминации [9, 11, 16].
При проникновении возбудителя со слюной клеща в макроорганизм, в области входных ворот может развиваться комплекс воспалительно-аллергических процессов, что проявляется эритемой, сопровождающейся регионарной лимфаденопатией. Множественные клинически значимые проявления ИКБ ассоциируются с наличием живых спирохет, которые связываются с поверхностью клеток человека, при активации которых запускается каскад выделения медиаторов воспаления [4, 5].
После размножения возбудителя в области входных ворот происходит лимфогенная и гематогенная диссеминация в ЛУ, внутренние органы, суставы, ЦНС. При этом наблюдается частичная гибель боррелий с высвобождением эндотоксина, обусловливающего явления интоксикации.
Генерализация инфекции клинически проявляется системным поражением внутренних органов - ЛУ, печени, мышц, сердечно-сосудистой и нервной системы, кожи. Боррелии способны проникать через гематоэнцефалический барьер, индуцируя соответствующую клиническую картину заболевания, а именно менингит, реже – менингоэнцефалит.
Способность боррелий к длительной персистенции в организме приводит к хроническому течению инфекционного процесса с поражением суставов и нервной системы.
Клинические проявления вне зависимости от стадии инфекционного процесса обусловлены наличием в организме живых боррелий и спирохетемией. Поражение различных систем организма в случае генерализации инфекции доказано результатами многочисленных экспериментов на животных, исследованиями прижизненных биоптатов органов и трупного материала [5, 9].
Отсутствие кожного процесса в дебюте, вероятно, сопряжено с трансмиссивным механизмом передачи инфекции, когда часть боррелий попадает непосредственно в кровяное русло [4, 11].
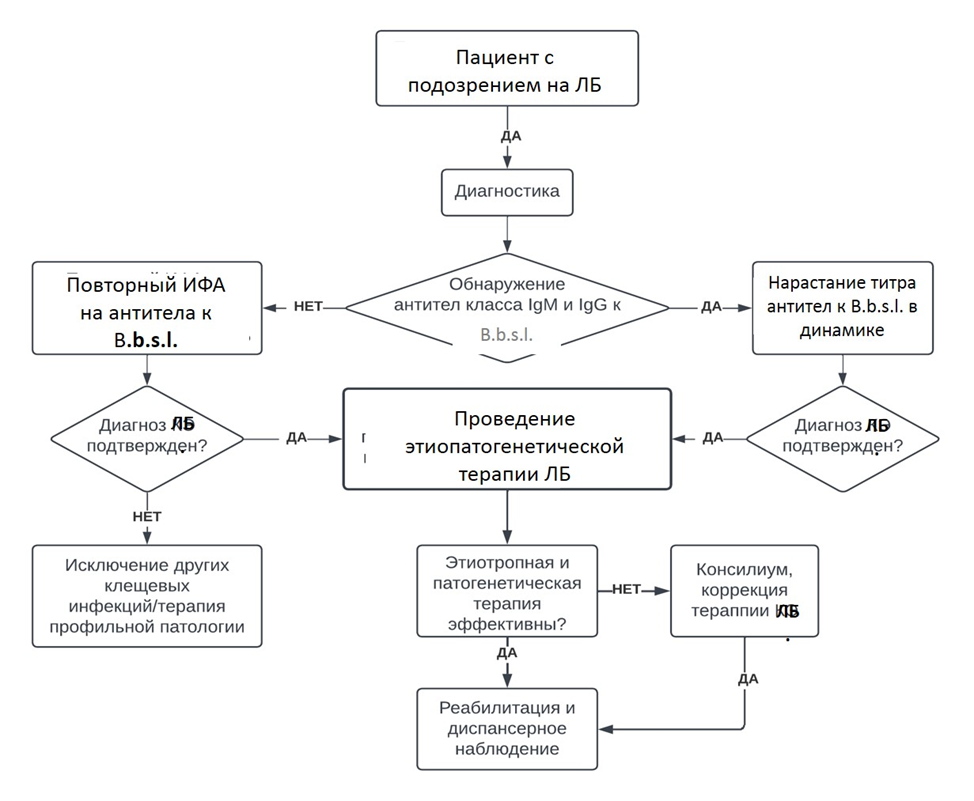
Специфический иммунный ответ при боррелиозной инфекции медленный. В ранние сроки заболевания вырабатываются специфические иммуноглобулины класса IgM, содержание которых достигает максимального уровня к 3-6й неделе болезни. Специфические антитела класса IgG обнаруживаются позднее - их концентрация увеличивается через 1,5-3-6 месяцев от начала заболевания [4, 5, 8, 17].
Боррелии способны взаимодействовать с множественными факторами специфической и неспецифической резистентности организма человека. В. Burgdorferi стимулируют выработку различных медиаторов воспаления (ИЛ-1, ИЛ-6, ФНО-α), участвующих в развитии артритов. В патогенезе нейроборрелиоза предполагается участие аутоиммунных реакций. Так, с помощью моноклональных антител обнаружены общие ангигенные детерминанты, в частности у флагеллярного антигена 41 kDа и некоторых тканей человека, а именно миелиновых волокон периферических нервов, нервных клетках и аксонах ЦНС, некоторых эпителиальных клетках, синусов и клетках синовиальных оболочек и кардиомиоцитах, т. е. именно в тех органах и тканях, которые наиболее часто поражаются при иксодовых клещевых боррелиозах. У больных острым боррелиозом уже в разгаре болезни повышается концентрация антител к денатурированной ДНК, митохондриям клеток, экстрагируемым ядерным антигенам и антител к компонентам ткани миокарда [4, 14, 17, 19].
На ранних стадиях фибробласты, макрофаги, эндотелиальные клетки и другие клетки организма продуцируют цитокины, состав которых в последующем может определить характер течения инфекции и её исхода. Именно в этот момент течения инфекционного процесса цитокины определяют пути дифференциации субтипов Т-клеток, обладающих патогенетическим потенциалом. [10, 11, 15, 16, 52, 53].
Активность Т-клеток, продуцирующих широкий спектр биологически активных веществ (БАВ), приводит к проявлению различных феноменов. Взаимодействие в этот момент иммуногенетических факторов, активация иммунокомпетентных клеток, формирование иммунного ответа и проявление аутоиммунных реакций приводит либо к элиминации возбудителя, либо к развитию различных клинико-патогенетических вариантов инфекции [10, 18].
Несомненно, ИКБ являются иммунозависимыми заболеваниями. Ответная реакция иммунной системы играет значимую роль в клиническом течении и исходах инфекционного процесса [5-7]. Незавершенный фагоцитоз в начальной стадии патогенеза, способность боррелий к длительной персистенции в фибробластах, выработка сериновой протеазы ВhpА обусловливают развитие рецидивирующих и хронических форм болезни [8, 17, 20]. Боррелии для защиты от иммунной реакции человека способны ингибировать эффект системы комплемента за счет поверхностных белков (Fhbp, OspE, OspF, Erps), связывающих плазматический регуляторный белок системы комплемента, усиливать синтез противовоспалительных цитокинов, связывать плазминоген с ОspA на своей поверхности, связываться с компонентами экстрацеллюлярного матрикса. Именно эти механизмы обусловливают способность боррелий к рекомбинационным перестройкам антигенной структуры поверхностных белков в процессе репродукции в организме человека, что способствует феномену «ускользания» от иммунного ответа [2, 5, 8, 16, 54].
Работами многих отечественных и зарубежных исследователей доказаны изменения различных звеньев иммунной системы при ИКБ [1, 2, 6, 8, 10, 12, 15, 16, 18, 55]. Реакция защиты начинается с фагоцитоза микроорганизмов и первичной воспалительной реакции. Активированные фагоциты секретируют цитокины с противовоспалительными (ИЛ-6, ИЛ-8, ФНО-а и пр.) и иммуностимулирующими (ИЛ-1, ИЛ-3, ИЛ-6, ИЛ-12, ИЛ-15, ФНО-а) эффектами.
ИЛ-1 участвует практически во всех этапах иммунного ответа. ИЛ-1 активирует АРС и СП4-лимфоциты, влияет на дифференцировку Т- и В-лимфоцитов и других иммунокомпетентных клеток. ИЛ-1 активирует цитотоксические СП8-лимфоциты и NK-клетки, участвует в регуляции продукции ИЛ-2, ИЛ-4, ИЛ-6, ИЛ- 8, гранулоцит-макрофаг колониестимулирующего фактора (GM-CSF) и других цитокинов. Активными ингибиторами продукции IL-1 являются IL-4, IL-10, IL- 12 и ФНО-а. ИЛ-3 — полипотентный активатор гемопоэтических клеток. Иммуностимулирующий эффект ИЛ-3 изучен недостаточно. Его реализация возможна через стимуляцию NK-клеток.
ИЛ-6 регулирует дифференцировку В-лимфоцитов и усиливает антителообразование, индуцирует цитотоксичность клеток. ИЛ-6 обладает выраженными провоспалительными действиями, принимает участие в генерации LAK-клеток и защищает нейтрофилы от апоптоза, усиливая их цитотоксический потенциал. ИЛ-6 усиливает синтез С-реактивного белка (СРБ). СРБ, после связывания с фосфолипидами клетки, активирует субкомпонент системы комплемента, запуская процесс, аналогичный классическому пути активации комплемента. Дальнейшее формирование мембраноатакующего комплекса заканчивается лизисом инфицированной клетки. Ингибирование воспаления интерлейконом-6 может быть связано и с его способностью индуцировать секрецию антагониста рецептора ИЛ-1р.
ИЛ-8 продуцируется многими типами клеток и обладает выраженными провоспалительными свойствами. Основным биологическим эффектом ИЛ-8 является индукция хемотаксиса нейтрофилов, эозинофилов, базофилов и других клеток системы иммунитета. IL-8 усиливает ангиогенез in vivo и in vitro. ИЛ-12 — полипотентный активатор клеточного иммунитета. Он усиливает активность Т-киллеров, NK- и LAK-клеток. ИЛ- 12 активирует цитотоксичность макрофагов. Важным свойством ИЛ-12 является усиление экспрессии FasL и индукция апоптоза. ИЛ-12 ингибирует ангиогенез. Антиангиогенное действие ИЛ-12 реализуется на уровне рецепторов протеинкиназ, адгезивных молекул, интегринов и других поверхностных структур.
ИЛ-15 по своим биологическим свойствам очень сходен с ИЛ-2 и во многом является его синергистом, в том числе, при индукции активности LAK-клеток. ИЛ-15 усиливает противовоспалительную активность Т- киллеров и NK-клеток, продукцию цитокинов CD4+ лимфоцитами и может проявлять себя как хемоаттрактант для Т-лимфоцитов.
Активация эндотелиоцитов сопровождается увеличением экспрессии на их мембранах белков типа ICAM- 1 И VCAM-1 и адгезией нейтрофилов, моноцитов и лимфоцитов. При сокращении активированного эндотелия межэндотелиальные щели расширяются, в них устремляются ранее фиксированные к эндотелию клетки; вокруг сосуда формируется первичное скопление моноцитов, которые превращаются в макрофаги будущей гранулемы. Макрофаги в очаге воспаления начинают вырабатывать медиаторы, привлекающие новые эффекторные клетки. Помимо цитокин-индуцирующей активации эндотелия, при воспалении возможна непосредственная его активация бактериальными клетками. Установлена способность поверхностных липопротеидов боррелий индуцировать экспрессию на активированном сосудистом эндотелии молекул адгезии — Е-селектинов, ICAM-1 И VCAM-1. Поверхностные липопротеиды боррелий также обладают хемотаксическими свойствами и способностью индуцировать секрецию монокинов.
Способность боррелий к самостоятельным поступательным движениям в тканях находит свое отражение в особенностях местного воспаления. В центре эритемы (в месте первоначального накопления возбудителя) боррелии подвергаются активному воздействию факторов воспаления, они теряют свою подвижность и их количество уменьшается, результатом чего является снижение явлений местного воспаления с формированием в центре эритемы просветления. Появление новых колец гиперемии связано с новыми генерациями боррелий за счет уменьшения факторов воспаления. Все это приводит обычно к развитию характерной мигрирующей эритемы, которая может и не иметь кольцевидную форму. Доказано, что при незавершенности фагоцитоза и неадекватности специфических гуморальных механизмов защиты формируются благоприятные условия для развития персистенции боррелий [15, 16]. Так же известно, что во все периоды болезни, по прошествии многих лет после заражения, можно обнаружить живых спирохет, в связи с чем лечение антибактериальными препаратами системного действия показано во всех стадиях. Этиотропное лечение в раннем периоде при мигрирующей эритеме способствует быстрому ее завершению и предупреждению развития поздних проявлений [5, 17].
В стадию диссеминации боррелий наблюдается гематогенное и лимфогенное распространение возбудителя из первичного очага в другие участки кожи и во внутренние органы. Не исключается способность боррелий к прямой пенетрации через эндотелиальные клетки в просвет сосудов. Распространение спирохет может происходить также лимфогенным путем и периневрально. Фиксированные боррелии вызывают гиперактивацию иммунной системы, формируется избыточный клеточный и гуморальный ответ; происходит активация Т-лимфоцитов с последующим повышением функциональной активности CDS-клеток, приводящим к истощению их популяции, системы комплемента и антител, состоянию относительного иммунодефицита [2, 4, 5, 7, 15]. Существенное значение имеют процессы, связанные с накоплением специфических иммунных комплексов, содержащих антигены боррелий, в синовиальной оболочке суставов, дерме, почках, миокарде.
Увеличение количества специфических иммуноглобулинов IgM наблюдается в период от 3 до 6 недели болезни, и связано с активацией В-лимфоцитов. Концентрация IgG в сыворотке крови повышается постепенно в течение месяцев, с увеличением числа боррелий и соответственно их антигенов. Лимфоциты и вырабатываемые ими специфические иммуноглобулины реагируют в начале заболевания избирательно на отдельные антигены боррелий, как поверхностные, так и цитоплазматические. По мере прогрессирования заболевания расширяется спектр антител к антигенам микробной клетки. Конечным этапом функционирования клеточного, гуморального звеньев иммунной системы и системы комплемента является образование комплексов антиген-антитело с последующей их элиминацией или лизисом. Длительно сохраняющийся высокий уровень специфических IgM у части больных при ИКБ некоторые исследователи связывают с поражением функции Т-хелперных клеток, необходимых для переключения продуцирования IgM на IgG, что может быть обусловлено применением ранней антибактериальной терапии, ослабляющей переключение синтеза антител [2, 4, 23-25].
В начале заболевания лимфоциты и вырабатываемые ими специфические иммуноглобулины реагируют избирательно на отдельные поверхностные и цитоплазматические антигены боррелий. Основными иммуногенами в патогенезе ИКБ являются поверхностные белки боррелий (липопротеин OspC). По мере прогрессирования заболевания расширяется спектр антител к антигенам микробной клетки (OspA, OspB). Эти липопротеины - мощные индукторы воспалительных реакций, что особенно характерно для OspA. Длительно сохраняясь на поверхности боррелий, они могут активировать как клетки неспецифической резистентности (макробактериофаги), так и поликлональные клетки 17 иммунной системы (В- и Т-лимфоциты). Активация поликлональных В-лимфоцитов приводит к синтезу антител, которые вызывают воспалительные реакции в соответствующем органе. При этом продуцируемые антитела обладают перекрестной иммунологической активностью как в отношении антигенов боррелий, так и тканевых белков (например, аксональные белки нервной ткани, белки синовиальной оболочки и др.). По мере прогрессирования заболевания аутоиммунные механизмы становятся все более значимы в патогенезе заболевания [4-6, 11, 15, 56].
Циркулирующие иммунные комплексы, образованные в результате взаимодействия антител и антигенов боррелий, подвергаются лизису или элиминации клетками СМФ. Увеличение количества ЦИК в крови приводит к накоплению их в синовиальной оболочке суставов, дерме, почках, миокарде. ЦИК привлекает в ткани нейтрофилы, вырабатывающие цитокины, биологически активные вещества и ферменты, вызывающие местные воспалительные и дистрофические изменения в пораженных тканях и органах.
Частым клиническим проявлением ИКБ на различных патогенетических стадиях является поражение опорно-двигательного аппарата: развитие артритов, тендинитов, бурситов, миозитов и пр. В патогенезе артритов основополагающую роль играют липопротеиды боррелий, стимулирующие секрецию ИЛ-1 клетками моноцитарно-макрофагального звена и цитотоксическими Т-лимфоцитами, в результате чего запускается синтез простагландинов и коллагеназы синовиальной тканью, и, как следствие, активация воспалительного процесса в суставах, резорбция костной и деструкция хрящевой ткани.
На стадии ранней диссеминации возможны элиминация боррелий и ЦИК, резидуальный метаморфоз патологического процесса и выздоровление.
До настоящего времени остается дискутабельным вопрос о механизмах длительной персистенции боррелии в организме больного. Хроническое течение болезни с поздними рецидивами и длительной (иногда более 10 лет) персистенцией боррелий в организме приводит к необратимым органным поражениям. В условиях иммунодефицитного состояния развиваются реактивно-аллергические реакции, активизируется аутоиммунный компонент воспаления, формируются стойкие дегенеративные и атрофические процессы в пораженных органах. Хроническое течение заболевания часто сопровождается поражением нервной, сердечно-сосудистой, опорно-двигательной систем.
Таким образом, со стороны макроорганизма причинами хронизации боррелиозной инфекции является неадекватный иммунный ответ: слабость механизмов формирования специфического иммунного ответа и склонность к образованию аутоантител к собственным тканям. Основными свойствами боррелий, способствующими их персистенции, являются значительная резистентность к воздействию системы комплемента и антигенная изменчивость.